
Specific Gene Expression in Pseudomonas Putida U Shows New Alternatives for Cadaverine and Putrescine Catabolism
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochemical Reagents
2.2. Microorganisms and Culture Conditions
2.3. Design of Primers for qPCR
2.4. Gene Induction and RNA Extraction
2.5. Standardizing qPCR
2.6. Validation of qPCR Amplicons
3. Results
3.1. Growth of P. putida U Using Putrescine, Cadaverine, GABA, DAVA, or Citrate as Carbon Sources
3.2. Search for Pseudomonas Putida U Homologous Genomes
3.3. Identification, Optimization, and Testing of qPCR Primers for SpuCs, GabTs, and Housekeeping Genes in P. putida U
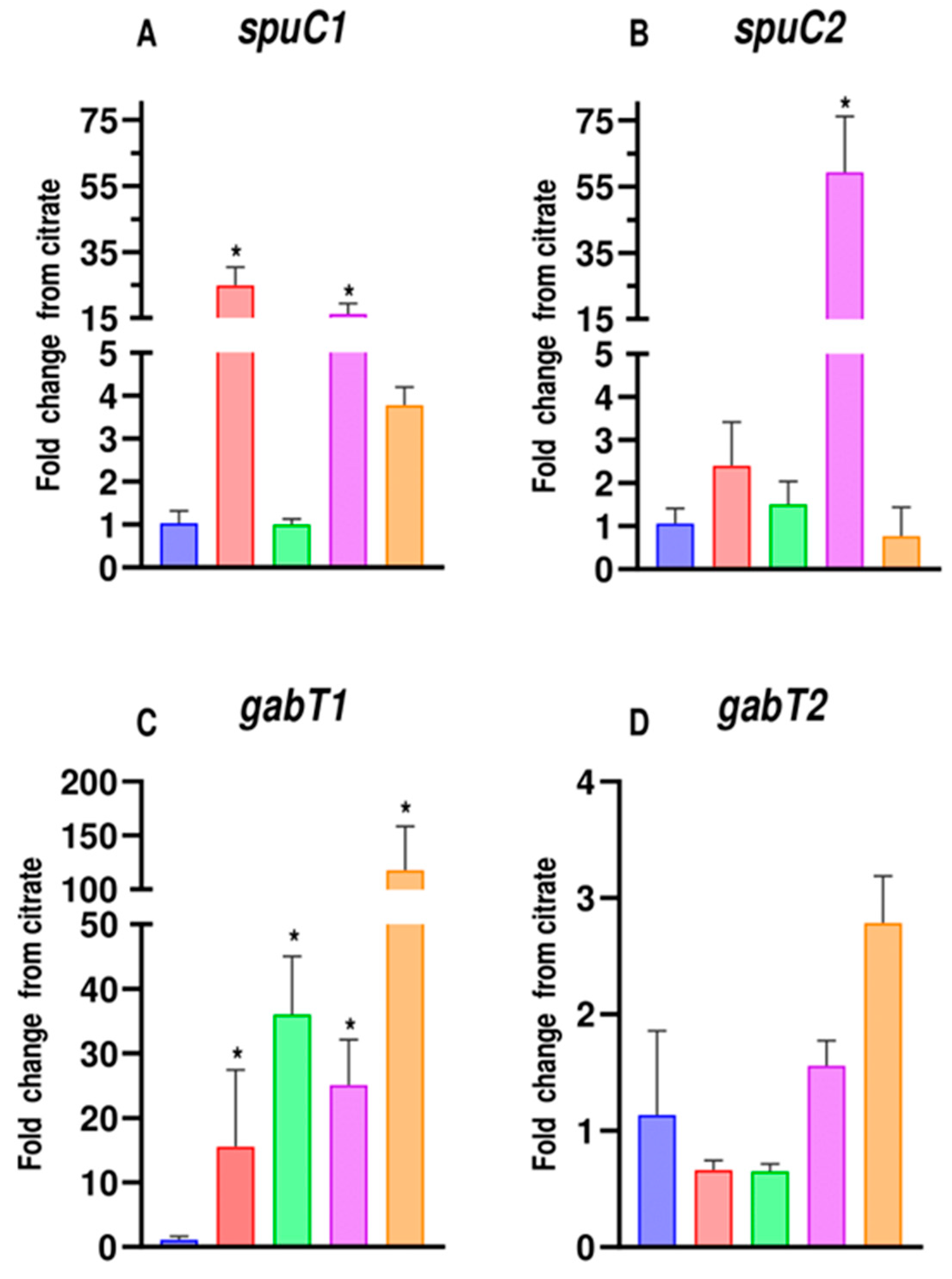
3.4. Expression Study of SpuCs and GabTs in Pseudomonas Putida U
3.5. Analysis of the Putative Cluster Containing SpuC1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yousefi, H.; Su, H.M.; Imani, S.M.; Alkhaldi, K.; Filipe, C.D.M.; Didar, T.F. Intelligent food packaging: A review of smart sensing technologies for monitoring food quality. ACS Sens. 2019, 4, 808–821. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of biogenic amines on food quality and safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Ovalle-Marmolejo, X.Y.; Redondo-Solano, M.; Granados-Chinchilla, F.; Miranda-Castilleja, D.E.; Arvizu-Medrano, S.M. Effect of stress factors on the production of biogenic amines by lactic acid bacteria isolated from fermented Mexican foods (cheese and beer). Food Control. 2023, 146, 109553. [Google Scholar] [CrossRef]
- Miguel, G.A.; Carlsen, S.; Arneborg, N.; Saerens, S.M.G.; Laulund, S.; Knudsen, G.M. Non-Saccharomyces yeasts for beer production: Insights into safety aspects and considerations. Int. J. Food Microbiol. 2022, 383, 109951. [Google Scholar] [CrossRef]
- Costantini, A.; Vaudano, E.; Pulcini, L.; Carafa, T.; Garcia-Moruno, E. An overview on biogenic amines in wine. Beverages 2019, 5, 19. [Google Scholar] [CrossRef]
- Moniente, M.; Botello-Morte, L.; Garcia-Gonzalo, D.; Pagan, R.; Ontanon, I. Analytical strategies for the determination of biogenic amines in dairy products. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3612–3646. [Google Scholar] [CrossRef] [PubMed]
- Schirone, M.; Esposito, L.; D’Onofrio, F.; Visciano, P.; Martuscelli, M.; Mastrocola, D.; Paparella, A. Biogenic amines in meat and meat products: A review of the science and future perspectives. Foods 2022, 11, 788. [Google Scholar] [CrossRef]
- Santos, M.H.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Gevrekci, A.Ö. The roles of polyamines in microorganisms. World J. Microbiol. Biotechnol. 2017, 33, 204. [Google Scholar] [CrossRef]
- Freudenberg, R.A.; Wittemeier, L.; Einhaus, A.; Baier, T.; Kruse, O. Advanced pathway engineering for phototrophic putrescine production. Plant Biotechnol. J. 2022, 20, 1968–1982. [Google Scholar] [CrossRef]
- Sagar, N.A.; Tarafdar, S.; Agarwal, S.; Tarafdar, A.; Sharma, S. Polyamines: Functions, metabolism, and role in human disease management. Med. Sci. 2021, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.S. A Guide to the Polyamines; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Young, C.C.; Chen, L.F. Polyamines in humic acid and their effect on radical growth of lettuce seedlings. Plant Soil 1997, 195, 143–149. [Google Scholar] [CrossRef]
- Wojcik, W.; Lukasiewicz, M.; Puppel, K. Biogenic amines: Formation, action and toxicity—A review. J. Sci. Food Agric. 2021, 101, 2634–2640. [Google Scholar] [CrossRef] [PubMed]
- Omer, A.K.; Mohammed, R.R.; Ameen, P.S.M.; Abas, Z.A.; Ekici, K. Presence of biogenic amines in food and their public health implications: A review. J. Food Prot. 2021, 84, 1539–1548. [Google Scholar] [CrossRef]
- Krysenko, S.; Wohlleben, W. Polyamine and ethanolamine metabolism in bacteria as an important component of nitrogen assimilation for survival and pathogenicity. Med. Sci. 2022, 10, 40. [Google Scholar] [CrossRef]
- Luengo, J.M.; Olivera, E.R. Catabolism of biogenic amines in Pseudomonas species. Environ. Microbiol. 2020, 22, 1174–1192. [Google Scholar] [CrossRef]
- Krysenko, S.; Matthews, A.; Busche, T.; Bera, A.; Wohlleben, W. Poly- and monoamine metabolism in Streptomyces coelicolor: The new role of glutamine synthetase-like enzymes in the survival under environmental stress. Microb. Physiol. 2021, 31, 233–247. [Google Scholar] [CrossRef]
- Ramakrishna, S.; Guarino, L.; Cohen, S.S. Polyamines of Anacystis nidulans and metabolism of exogenous spermidine and spermine. J. Bacteriol. 1978, 134, 744–750. [Google Scholar] [CrossRef]
- Samsonova, N.N.; Smirnov, S.V.; Altman, I.B.; Ptitsyn, L.R. Molecular cloning and characterization of Escherichia coli K12 ygjG gene. BMC Microbiol. 2003, 3, 2. [Google Scholar] [CrossRef]
- Samsonova, N.N.; Smirnov, S.V.; Novikova, A.E.; Ptitsyn, L.R. Identification of Escherichia coli K12 YdcW protein as a γ-aminobutyraldehyde dehydrogenase. FEBS Lett. 2005, 579, 4107–4112. [Google Scholar] [CrossRef]
- Kurihara, S.; Oda, S.; Kato, K.; Kim, H.G.; Koyanagi, T.; Kumagai, H.; Suzuki, H. A novel putrescine utilization pathway involves γ-glutamylated intermediates of Escherichia coli K-12. J. Biol. Chem. 2005, 280, 4602–4608. [Google Scholar] [CrossRef]
- Suzuki, H.; Kurihara, S. Polyamine catabolism in prokaryotes. In Polyamines; Springer: Tokyo, Japan, 2015; pp. 47–59. [Google Scholar]
- Schneider, B.L.; Reitzer, L. Pathway and enzyme redundancy in putrescine catabolism in Escherichia coli. J. Bacteriol. 2012, 194, 4080–4088. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, S.; Kato, K.; Asada, K.; Kumagai, H.; Suzuki, H. A putrescine-inducible pathway comprising PuuE-YneI in which γ-aminobutyrate is degraded into succinate in Escherichia coli K-12. J. Bacteriol. 2010, 192, 4582–4591. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.D.; Itoh, Y.; Nakada, Y.; Jiang, Y. Functional analysis and regulation of the divergent spuABCDEFGH-spuI operons for polyamine uptake and utilization in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2002, 184, 3765–3773. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.T.; Kwon, D.H.; Hegazy, M.; Lu, C.D. Transcriptome analysis of agmatine and putrescine catabolism in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2008, 190, 1966–1975. [Google Scholar] [CrossRef]
- Yao, X.; He, W.; Lu, C.-D. Functional characterization of seven γ-glutamylpolyamine synthetase genes and the bauRABCD locus for polyamine and β-alanine utilization in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2011, 193, 3923–3930. [Google Scholar] [CrossRef]
- Chou, H.T.; Li, J.Y.; Peng, Y.C.; Lu, C.D. Molecular characterization of PauR and its role in control of putrescine and cadaverine catabolism through the γ-glutamylation pathway in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2013, 195, 3906–3913. [Google Scholar] [CrossRef]
- Arcos, M.; Olivera, E.R.; Arias, S.; Naharro, G.; Luengo, J.M. The 3,4-dihydroxyphenylacetic acid catabolon, a catabolic unit for degradation of biogenic amines tyramine and dopamine in Pseudomonas putida U. Environ. Microbiol. 2010, 12, 1684–1704. [Google Scholar] [CrossRef]
- Arias, S.; Olivera, E.R.; Arcos, M.; Naharro, G.; Luengo, J.M. Genetic analyses and molecular characterization of the pathways involved in the conversion of 2-phenylethylamine and 2-phenylethanol into phenylacetic acid in Pseudomonas putida U. Environ. Microbiol. 2008, 10, 413–432. [Google Scholar] [CrossRef]
- de la Torre, M.; Gómez-Botrán, J.L.; Olivera, E.R.; Bermejo, F.; Rodríguez-Morán, J.; Luengo, J.M. Histamine catabolism in Pseudomonas putida U: Identification of the genes, catabolic enzymes and regulators. Environ. Microbiol. 2018, 20, 1828–1841. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis I. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Blanco, H.; Reglero, A.; Rodriguez-Aparicio, L.B.; Luengo, J.M. Purification and biochemical characterization of phenylacetyl-CoA ligase from Pseudomonas putida. A specific enzyme for the catabolism of phenylacetic acid. J. Biol. Chem. 1990, 265, 7084–7090. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.Z.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Saitou, N. Property and efficiency of the maximum likelihood method for molecular phylogeny. J. Mol. Evol. 1988, 27, 261–273. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Cardoza, R.E.; McCormick, S.P.; Lindo, L.; Kim, H.S.; Olivera, E.R.; Nelson, D.R.; Proctor, R.H.; Gutierrez, S. A cytochrome P450 monooxygenase gene required for biosynthesis of the trichothecene toxin harzianum A in Trichoderma. Appl. Microbiol. Biotechnol. 2019, 103, 8087–8103. [Google Scholar] [CrossRef] [PubMed]
- Lindo, L.; Cardoza, R.E.; Lorenzana, A.; Casquero, P.A.; Gutierrez, S. Identification of plant genes putatively involved in the perception of fungal ergosterol-squalene. J. Integr. Plant Biol. 2020, 62, 927–947. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.; Lood, C.; Rokni-Zadeh, H.; van Noort, V.; Lavigne, R.; De Mot, R. Reliable identification of environmental Pseudomonas isolates using the rpoD gene. Microorganisms 2020, 8, 1166. [Google Scholar] [CrossRef] [PubMed]
- Pazos-Rojas, L.A.; Munoz-Arenas, L.C.; Rodriguez-Andrade, O.; Lopez-Cruz, L.E.; Lopez-Ortega, O.; Lopes-Olivares, F.; Luna-Suarez, S.; Baez, A.; Morales-Garcia, Y.E.; Quintero-Hernandez, V.; et al. Desiccation-induced viable but nonculturable state in Pseudomonas putida KT2440, a survival strategy. PLoS ONE 2019, 14, e0219554. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Galman, J.L.; Gahloth, D.; Parmeggiani, F.; Slabu, I.; Leys, D.; Turner, N.J. Characterization of a putrescine transaminase from Pseudomonas putida and its application to the synthesis of benzylamine derivatives. Front. Bioeng. Biotechnol. 2018, 6, 205. [Google Scholar] [CrossRef]
- Hasne, M.P.; Coppens, I.; Soysa, R.; Ullman, B. A high-affinity putrescine-cadaverine transporter from Trypanosoma cruzi. Mol. Microbiol. 2010, 76, 78–91. [Google Scholar] [CrossRef]
- Kroger, P.; Shanmugaratnam, S.; Ferruz, N.; Schweimer, K.; Hocker, B. A comprehensive binding study illustrates ligand recognition in the periplasmic binding protein PotF. Structure 2021, 29, 433.e4. [Google Scholar] [CrossRef]
- Szalai, G.; Janda, K.; Darko, E.; Janda, T.; Peeva, V.; Pal, M. Comparative analysis of polyamine metabolism in wheat and maize plants. Plant Physiol. Biochem. 2017, 112, 239–250. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/review: Contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci. 2012, 193, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Galman, J.L.; Slabu, I.; Weise, N.J.; Iglesias, C.; Parmeggiani, F.; Lloyd, R.C.; Turner, N.J. Biocatalytic transamination with near-stoichiometric inexpensive amine donors mediated by bifunctional mono- and di-amine transaminases. Green Chem. 2017, 19, 361–366. [Google Scholar] [CrossRef]
- Yu, P.; Ren, Q.; Wang, X.X.; Huang, X.X. Enhanced biosynthesis of gamma-aminobutyric acid (GABA) in Escherichia coli by pathway engineering. Biochem. Eng. J. 2019, 141, 252–258. [Google Scholar] [CrossRef]
- Arias-Barrau, E.; Sandoval, A.; Naharro, G.; Olivera, E.R.; Luengo, J.M. A two-component hydroxylase involved in the assimilation of 3-hydroxyphenyl acetate in Pseudomonas putida. J. Biol. Chem. 2005, 280, 26435–26447. [Google Scholar] [CrossRef]
- Merino, E.; Barrientos, A.; Rodriguez, J.; Naharro, G.; Luengo, J.M.; Olivera, E.R. Isolation of cholesterol- and deoxycholate-degrading bacteria from soil samples: Evidence of a common pathway. Appl. Microbiol. Biotechnol. 2013, 97, 891–904. [Google Scholar] [CrossRef]
- Olivera, E.R.; Minambres, B.; Garcia, B.; Muniz, C.; Moreno, M.A.; Ferrandez, A.; Diaz, E.; Garcia, J.L.; Luengo, J.M. Molecular characterization of the phenylacetic acid catabolic pathway in Pseudomonas putida U: The phenylacetyl-CoA catabolon. Proc. Natl. Acad. Sci. USA 1998, 95, 6419–6424. [Google Scholar] [CrossRef]
- Garcia, B.; Olivera, E.R.; Minambres, B.; Fernandez-Valverde, M.; Canedo, L.M.; Prieto, M.A.; Garcia, J.L.; Martinez, M.; Luengo, J.M. Novel biodegradable aromatic plastics from a bacterial source: Genetic and biochemical studies on a route of the phenylacetyl-CoA catabolon. J. Biol. Chem. 1999, 274, 29228–29241. [Google Scholar] [CrossRef]
- Arias-Barrau, E.; Olivera, E.R.; Luengo, J.M.; Fernandez, C.; Galan, B.; Garcia, J.L.; Diaz, E.; Minambres, B. The homogentisate pathway: A central catabolic pathway involved in the degradation of L-phenylalanine, L-tyrosine, and 3-hydroxyphenylacetate in Pseudomonas putida. J. Bacteriol. 2004, 186, 5062–5077. [Google Scholar] [CrossRef]
- Olivera, E.R.; Carnicero, D.; Garcia, B.; Minambres, B.; Moreno, M.A.; Canedo, L.; DiRusso, C.C.; Naharro, G.; Luengo, J.M. Two different pathways are involved in the beta-oxidation of n-alkanoic and n-phenylalkanoic acids in Pseudomonas putida U: Genetic studies and biotechnological applications. Mol. Microbiol. 2001, 39, 863–874. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide | Target Gene | Sequence (5’–3’) | Amplicon Size (bp) | Efficiency (%) | R2 of Efficiencies |
|---|---|---|---|---|---|
| Q-SpuC1,1-5′ PPU | spuC1 | AAGCCGCACAAGCAGACCAT | 132 pb | 96.59 | 0.990 |
| Q-SpuC1,1-3′ PPU | GATGTGCACGATGCCCGGAA | ||||
| Q-SpuC2,1-5′ PPU | spuC2 | CCAAGGGCGTGTACCTGTGG | 122 pb | 95.20 | 0.986 |
| Q-SpuC2,1-3′ PPU | CATCTGCTGGCTGGCAACCT | ||||
| F1.gabT1.PPU | gabT1 | GCTGTTGGCCAGGTTGTTGC | 144 pb | 98.82 | 0.998 |
| R1.gabT1.PPU | CAGTGCGTCTTCGGCAGTCA | ||||
| F1.gabT2.PPU | gabT2 | CGCCATCGTTCACCCGATCA | 120 pb | 93.92 | 0.992 |
| R1.gabT2.PPU | TTGCAGTGGCCCAGGTTGAG | ||||
| F1.rpoD.PPU | rpoD | GCGAACCTGCGTCTGGTGAT | 138 pb | 96.14 | 0.997 |
| R1.rpoD.PPU | GAACTTGTAGCCGCGACGGT | ||||
| F1.rpoN.PPU | rpoN | TCGAACCCGATGCTCGAACG | 109 pb | 101.01 | 0.994 |
| R1.rpoN.PPU | AGCTGTTGTCCTGGGCTTCG | ||||
| F1.recA.PPU | recA | GGCGAACAGGCCCTGGAAAT | 107 pb | 99.42 | 0.988 |
| R1.recA.PPU | CCTTCGATCTCGGCCTTGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Getino, L.; Chamizo-Ampudia, A.; Martín, J.L.; Luengo, J.M.; Barreiro, C.; Olivera, E.R. Specific Gene Expression in Pseudomonas Putida U Shows New Alternatives for Cadaverine and Putrescine Catabolism. Genes 2023, 14, 1897. https://doi.org/10.3390/genes14101897
Getino L, Chamizo-Ampudia A, Martín JL, Luengo JM, Barreiro C, Olivera ER. Specific Gene Expression in Pseudomonas Putida U Shows New Alternatives for Cadaverine and Putrescine Catabolism. Genes. 2023; 14(10):1897. https://doi.org/10.3390/genes14101897
Chicago/Turabian StyleGetino, Luis, Alejandro Chamizo-Ampudia, José Luis Martín, José María Luengo, Carlos Barreiro, and Elías R. Olivera. 2023. "Specific Gene Expression in Pseudomonas Putida U Shows New Alternatives for Cadaverine and Putrescine Catabolism" Genes 14, no. 10: 1897. https://doi.org/10.3390/genes14101897